




中华医学会
地址: 中国北京东四西大街42号
邮编: 100710
缺氧诱导因子1α调控缺氧诱导的新生大鼠肺动脉内皮细胞增殖机制研究
摘要
目的
探讨缺氧诱导因子1α(hypoxia induced factor-1α,HIF-1α)通过丙酮酸脱氢酶激酶1(pyruvate dehydrogenase kinase 1,PDK1)调控缺氧诱导的新生大鼠肺动脉内皮细胞(pulmonary artery endothelial cells,PAECs)增殖的作用机制。
方法
使用36只体重20~30 g、日龄7~10 d 的Wistar新生大鼠,对其PAECs进行分离、培养后,将体外培养的PAECs随机分为常氧组、缺氧组、HIF-1α+常氧组、HIF-1α+缺氧组、HIF-1α抑制剂(2-ME)+缺氧组、PDK1抑制剂+缺氧组。HIF-1α+常氧组和HIF-1α+缺氧组以腺病毒转染HIF-1α。于24 h、48 h和72 h,应用CCK8检测PAECs增殖能力,伤口愈合实验检测PAECs迁移率,实时荧光定量聚合酶链式反应及Western blotting实验检测PAECs中HIF-1α和PDK1的mRNA及蛋白表达水平,相应试剂盒检测PAECs葡萄糖摄取量及乳酸生成量。
结果
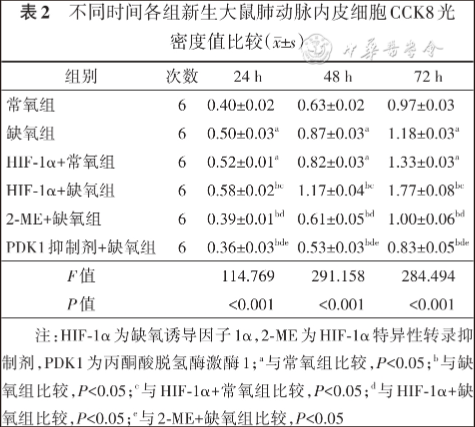
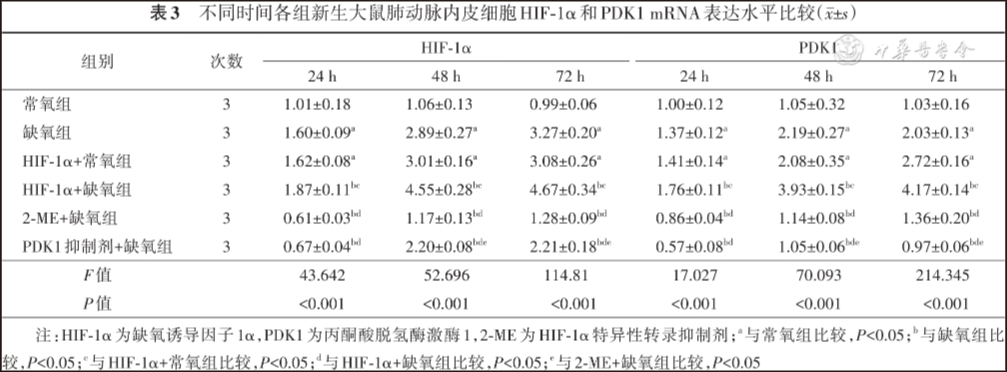
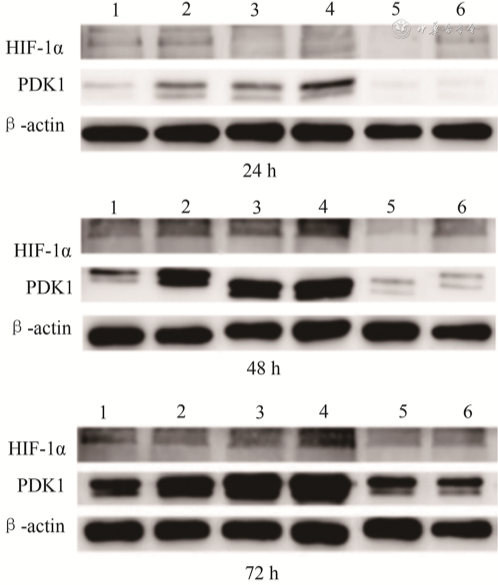
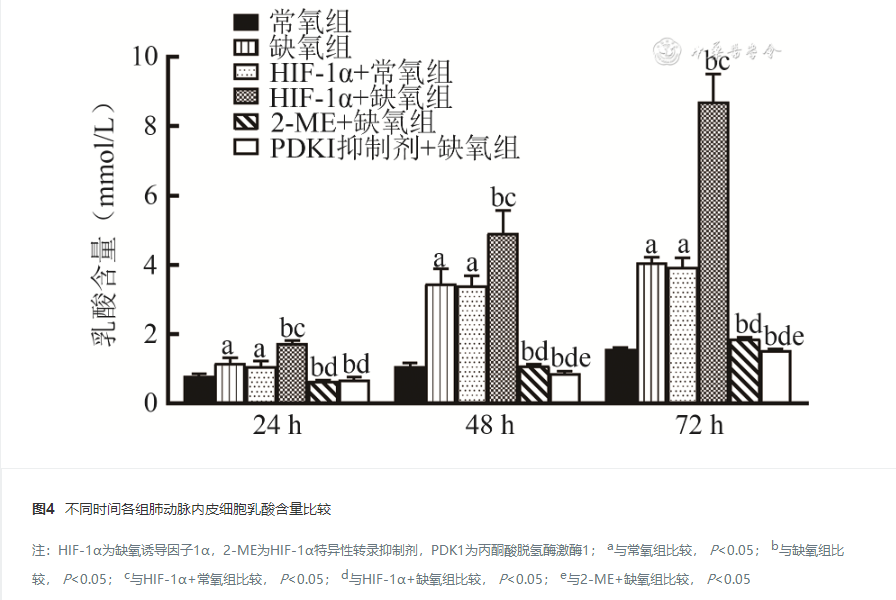
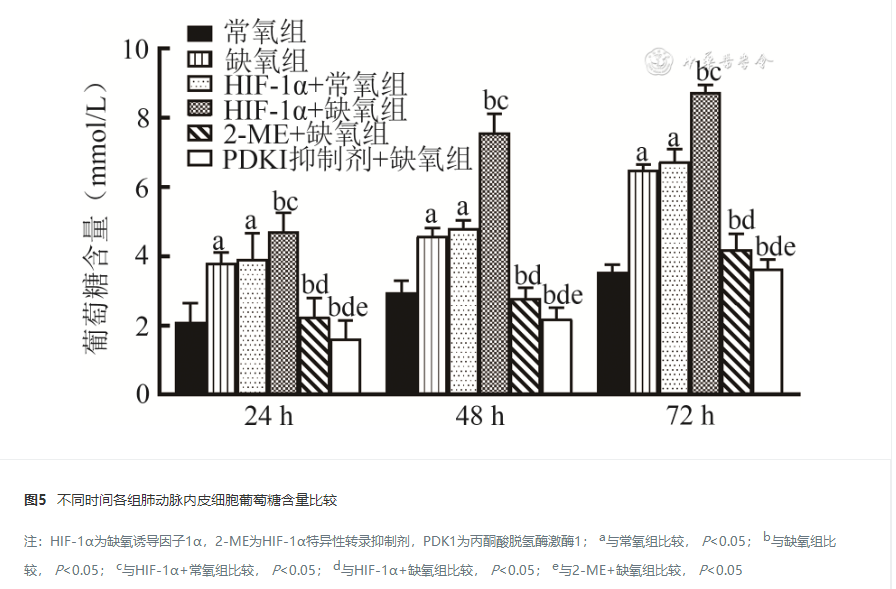
24 h、48 h及72 h,缺氧组和HIF-1α+常氧组PAECs增殖及迁移能力、HIF-1α和PDK1蛋白及mRNA表达水平、乳酸生成及葡萄糖摄入量高于常氧组,HIF-1α+缺氧组PAECs增殖及迁移能力、HIF-1α和PDK1蛋白及mRNA表达水平、乳酸生成及葡萄糖摄入量高于其他缺氧组,PDK1抑制剂+缺氧组和2-ME+缺氧组PAECs增殖及迁移能力、HIF-1α和PDK1蛋白及mRNA表达水平、乳酸生成及葡萄糖摄取量低于缺氧组,差异均有统计学意义( P<0.05)。
结论
缺氧条件下,HIF-1α可能通过上调PDK1的表达增强Warburg效应,促进新生大鼠PAECs的增殖。
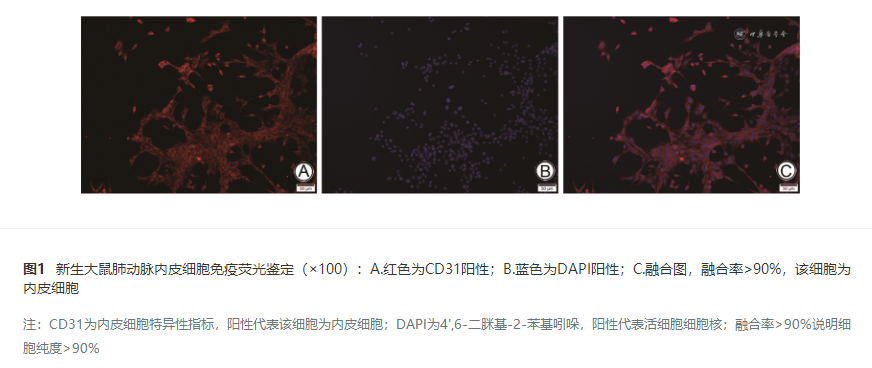
新生儿缺氧性肺动脉高压(hypoxic pulmonary hypertension,HPH)是新生儿重症监护室常见的急危重症 [ 1 , 2 ] ,尚无有效治疗方法。其病理特征为肺血管阻力和血管重塑的不断升级,发病机制尚未明确 [ 3 ] 。肺动脉内皮细胞(pulmonary artery endothelial cells,PAECs)在血管重塑过程中起重要作用 [ 4 , 5 , 6 ] 。PAECs增殖机制中存在糖代谢通路异常 [ 7 ] ,在氧气充足的情况下以高糖吸收、有氧糖酵解及高乳酸生成为主,即Warburg效应 [ 8 ] ,丙酮酸脱氢酶激酶1(pyruvate dehydrogenase kinase 1,PDK1)是通过磷酸化负调节丙酮酸脱氢酶复合物(pyruvate dehydrogenase complex,PDC)活性的关键酶,而PDC失活与Warburg效应紧密相关 [ 9 ] ,并且PDK1可能参与缺氧期间内皮细胞增殖和凋亡的调节 [ 10 ] 。 在缺氧条件下,缺氧诱导因子1α(hypoxia induced factor-1 alpha,HIF-1α)与缺氧反应元件(hypoxia response elements,HRE)结合激活缺氧介导反应,转录并活化下游靶基因调控糖酵解的PDK1 [ 11 , 12 , 13 ] ,HIF-1α还可调控下游靶基因引起肺血管痉挛及重塑,造成新生大鼠肺动脉高压 [ 14 ] ,但HIF-1α是否通过PDK1影响Warburg效应参与HPH肺血管重塑过程尚未知晓。本研究旨在探讨HIF-1α是否促进PDK1表达增强Warburg效应,从而导致缺氧PAECs过度增殖,为气道重塑机制提供新的见解,并为HPH的治疗和潜在药物靶点提供实验依据。 材料和方法 一、实验动物及主要材料 1.实验动物:选择体重20~30 g、日龄7~10 d的Wistar新生大鼠36只,雌雄比例1∶1,购自新疆医科大学动物实验中心,动物许可证号:SCXK(新)2011-0004。所有动物均饲养于25℃、55%湿度、12 h明暗交替环境,保证充足的食物和水。本实验已通过新疆医科大学第一附属医院伦理委员会审批(K202409-13)。 2.主要试剂与仪器:便携式细胞低氧培养箱(美国billups-rothenberg公司)、CY-100B氧浓度控制仪(上海玉研科学仪器有限公司);超净工作台(上海精密仪器仪表有限公司);解剖显微镜(南京雷炯仪器设备有限公司)、BX41TF光学显微镜(日本奥林巴斯)、相差显微镜(中科美菱低温科技有限责任公司);HIF-1α腺病毒(上海GeneChem)、HIF-1α特异性转录抑制剂2-ME(广州威佳科技有限公司)、PDK1特异性抑制剂(美国APE x BIO)、乳酸和葡萄糖检测试剂盒(北京索莱宝科技有限公司)。 二、实验方法 1.原代PAECs分离:所有Wistar新生大鼠断颈处死,75%酒精浸泡后取出肺动脉,将动脉内膜面贴入培养皿中并加入消化液,恒温培养箱中孵育后,用显微镊刮取肺动脉内膜面,吹打消化液并收集,细胞筛过滤后离心、弃上清,保留细胞沉淀。 2.原代PAECs鉴定:将细胞爬片放入24孔板中,注入内皮细胞完全培养基,放入37℃、5%CO 2恒温培养箱中培养贴壁后进行固定,室温通透、封闭,CD31(PAECs的特异性指标)孵育过夜,荧光(Cy3)标记羊抗小鼠IGg后湿盒孵育,4',6-二脒基-2-苯基吲哚(4',6-diamidino-2-phenylindole,DAPI)(代表活细胞核)避光孵育,封片后荧光显微镜下观察并采集图片。 3.原代PAECs培养:内皮细胞完全培养基重悬细胞后接种于培养皿中,37℃、5%CO 2恒温培养箱中静置培养,48 h后更换新鲜培养基,以后3 d换液一次,细胞长满后使用。 4.实验分组及缺氧PAECs模型建立:将PAECs按照单纯随机分组方法分为常氧组、缺氧组、HIF-1α+常氧组(HIF-1α腺病毒转染并常氧培养)、HIF-1α+缺氧组(HIF-1α腺病毒转染并缺氧培养)、2-ME+缺氧组(HIF-1α抑制剂干预并缺氧培养)、PDK1抑制剂+缺氧组(PDK1抑制剂干预并缺氧培养)。以上各组根据观察时间点另分为24 h、48 h和72 h亚组。常氧组细胞于37℃、5%CO 2恒温培养箱中培养,缺氧组细胞移至缺氧小室内,灌注由3%O 2、5%CO 2、92%N 2组成的混合气体后密封,置于37℃恒温培养箱中培养。 5.HIF-1α腺病毒转染:将生长状态良好的细胞制备成细胞悬液接种于6孔板内,待细胞融合至60%,吸弃培养液,加入含有HIF-1α的腺病毒(标有绿色荧光蛋白并携带HIF-1α基因的腺病毒载体)、病毒增强液、完全培养基的混合液(腺病毒滴度为1×10 10,最适感染复数值为100,病毒和增强液按照1∶1比例进行转染),8~12 h后更换为完全培养基。于转染24 h、48 h及72 h后在倒置荧光显微镜下观察腺病毒转染情况。 6.HIF-1α及PDK1抑制:HIF-1α抑制剂2-ME和PDK1抑制剂均以粉末形态存在,能在蛋白水平上抑制这两种物质的表达,使用二甲基亚砜作为溶剂。对目标细胞进行饥饿同步化处理后,再用培养基孵育12 h,随后将溶解好2-ME和PDK1抑制剂的二甲基亚砜溶液与完全培养基混合,用于细胞培养。在24 h、48 h、72 h对处理过的细胞进行后续实验操作。 7.CCK8实验检测细胞活性与增殖:用CCK8法检测PAECs活性与增殖,波长450 nm处的光密度值与细胞数量呈正相关。生长状态良好的PAECs以每孔细胞数5 000接种于96孔板中,根据分组干预处理后继续培养,于各检测时间将培养液更换为100 μl的新鲜完全培养基,并加入10 μl CCK8试剂,避光置于培养箱内孵育2 h后使用分光光度计测定光密度值。 8.伤口愈合实验测定细胞迁移:将生长状态良好的PAECs接种至6孔板中,密度达80%~90%时进行纵向划痕,洗去漂浮细胞后加入低血清培养基并进行干预处理。于划痕0 h、24 h、48 h和72 h选取同一视野使用显微镜采集图片,使用Image-Pro plus6.0软件测量划痕面积。迁移率计算公式:迁移率=(0 h划痕面积-该时间点划痕面积)/0 h划痕面积。 9.实时荧光定量聚合酶链式反应(polymerase chain reaction,PCR)检测PAECs中目的基因mRNA表达情况:弃去培养瓶内培养液,加入Buffer cRLQ消化裂解细胞,Trizol法提取RNA,分光光度计检测RNA浓度及纯度,冰上配置10 μl逆转录反应体系。上下游引物由上海生物工程合成有限公司合成,结果通过2-ΔΔCt法分析并计算。引物序列见 表1 。 10.Western Blotting实验检测PAECs内目的基因蛋白表达情况:弃去培养瓶内培养液,加入RIPA裂解液,提取总蛋白后,蛋白样品经十二烷基硫酸钠-聚丙烯酰胺凝胶电泳,转至PVDF膜,封闭后分别加入HIF-1α(1∶1 000)、PDK1(1∶1 000)、GAPDH(1∶1 000)抗体,4℃孵育过夜,加入二抗(1∶1 000),室温孵育2 h,加入增强型化学发光试剂显影,采用ImageJ对条带进行灰度值分析。 11.乳酸含量检测:PAECs用胰蛋白酶消化重悬后以10 5个/孔接种于6孔板内,各组干预处理后继续培养,不更换培养基,于24 h、48 h、72 h收集细胞培养基上清液,按照说明加入提取液,离心取上清,与标准品分别加入各反应液后37℃避光反应20 min,离心后取沉淀,乙醇充分溶解后在酶标仪570 nm处测定吸光值,根据标准品绘制出的标准曲线和对应公式计算乳酸含量。 12.葡萄糖含量检测:PAECs用胰蛋白酶消化重悬后以10 5个/孔接种于6孔板内,各组干预处理后继续培养,不更换培养基,于24 h、48 h、72 h收集细胞培养基上清液,按照说明于1.5 ml EP管中加入各反应试剂,涡旋混匀,100℃煮沸15 min,冷却至室温后,取200 μl至96孔板中测定630 nm处吸光值,按照对应公式计算葡萄糖含量。 三、统计学方法 应用SPSS 26.0统计软件对数据进行分析,应用Graphpad Prism 8.0进行制图。计量资料均进行Shapiro-Wilk正态性检验,符合正态分布的计量资料以均值±标准差表示,固定时间因素,多组间比较采用单因素方差分析(One-way ANOVA),组间两两比较采用LSD检验。 P<0.05为差异有统计学意义。 结 果 一、PAECs细胞鉴定情况 PAECs阳性率>90%,即细胞纯度>90%,见 图1 。 二、各组PAECs活性与增殖能力 24 h、48 h、72 h各组PAECs活性与增殖能力比较,缺氧组和HIF-1α+常氧组高于常氧组,HIF-1α+缺氧组高于缺氧组、2-ME+缺氧组及PDK1抑制剂+缺氧组,2-ME+缺氧组和PDK1抑制剂+缺氧组低于缺氧组,差异均有统计学意义( P<0.05)。见 表2 。 三、各组PAECs迁移率 24 h、48 h、72 h各组迁移率比较,缺氧组和HIF-1α+常氧组高于常氧组,HIF-1α+缺氧组高于缺氧组、2-ME+缺氧组、PDK1抑制剂+缺氧组,2-ME+缺氧组和PDK1抑制剂+缺氧组低于缺氧组,差异均有统计学意义( P<0.05)。见 图2 。 四、各组大鼠PAECs中HIF-1α和PDK1 mRNA表达水平 24 h、48 h、72 h各组HIF-1α及PDK1 mRNA表达水平比较,缺氧组和HIF-1α+常氧组高于常氧组,HIF-1α+缺氧组高于缺氧组、2-ME+缺氧组、PDK1抑制剂+缺氧组,2-ME+缺氧组和PDK1抑制剂+缺氧组低于缺氧组,差异均有统计学意义( P<0.05)。见 表3 。 五、各组大鼠PAECs中HIF-1α和PDK1蛋白表达水平 24 h、48 h、72 h各组HIF-1α和PDK1蛋白表达水平比较,缺氧组和HIF-1α+常氧组高于常氧组,HIF-1α+缺氧组高于缺氧组、2-ME+缺氧组、PDK1抑制剂+缺氧组,2-ME+缺氧组和PDK1抑制剂+缺氧组低于缺氧组,差异均有统计学意义( P<0.05)。见 图3 。 六、各组大鼠PAECs乳酸及葡萄糖含量 24 h、48 h、72 h各组乳酸及葡萄糖含量比较,缺氧组和HIF-1α+常氧组高于常氧组,HIF-1α+缺氧组高于缺氧组、2-ME+缺氧组、PDK1抑制剂+缺氧组,2-ME+缺氧组和PDK1抑制剂+缺氧组低于缺氧组,差异均有统计学意义( P<0.05)。见 图4 、 5 。 讨 论 新生儿HPH是由血管收缩和肺血管重塑引起的肺动脉压升高以及右心室肥厚 [ 15 ] ,致死率高,确切机制尚未完全明了 [ 16 ] 。肺血管重塑为HPH的重要特征,涉及PAECs及肺动脉平滑肌细胞的增殖和迁移失调,其中内皮功能异常在介导肺血管系统结构变化中起至关重要的作用 [ 17 ] ,在肺动脉高压早期通过改善PAECs功能障碍促进血管重塑为目前研究的关注点 [ 18 , 19 ] 。而PAECs增殖严重依赖糖酵解 [ 20 ] ,研究显示,成人肺动脉高压和右心室衰竭患者PDK1表达增加 [ 21 ] ,沉默PDK1可抑制缺氧诱导的肺动脉高压 [ 22 ] 。本研究结果显示,在各观察时间点,缺氧组的PAECs增殖能力、迁移率、乳酸及葡萄糖含量均高于常氧组,且PDK1蛋白及mRNA表达水平高于常氧组。PDK1抑制剂+缺氧组PAECs的增殖及迁移能力、PDK1蛋白及mRNA表达量、乳酸和葡萄糖含量低于缺氧组。说明缺氧环境促进PAECs PDK1表达,增强PAECs的Warburg效应,刺激其增殖和迁移,使用PDK1特异性转录抑制剂后,PAECs的Warburg效应随之减弱,细胞增殖和迁移能力受抑制,提示缺氧对PAECs异常增殖的作用机制与PDK1激活及Warburg效应活跃有关。 HIF-1α是细胞适应缺氧的关键调节因子,缺氧条件下HIF-1α被激活并促进一系列表达变化,包括参与细胞代谢、血管生成、细胞增殖 [ 23 ] 。临床研究显示,HPH新生儿肺动脉收缩压水平与血清HIF-1α水平成正相关 [ 24 ] ,HIF-1α表达上调可启动HPH的血管重塑 [ 25 ] ,并且参与肺血管重塑过程中PAECs的过度增殖和抗凋亡 [ 26 ] 。本课题组前期研究也发现HPH新生大鼠HIF-1α水平升高 [ 27 , 28 ] ,HIF-1α与PDK1可能与HPH的发展有关。为验证HIF-1α与PDK1在HPH中的作用机制,本研究通过转染含有HIF-1α的腺病毒载体实现胞内HIF-1α过表达,并观察到标记腺病毒的绿色荧光,表明转染成功。 本研究结果显示,缺氧组和HIF-1α+常氧组PAECs的HIF-1α蛋白及mRNA表达均较常氧组明显升高,HIF-1α+缺氧组较缺氧组明显升高;PAECs增殖和迁移能力结果显示,缺氧组和HIF-1α+缺氧组高于缺氧组,HIF-1α+常氧组高于常氧组,其中HIF-1α+缺氧组最高,提示缺氧促进PAECs中HIF-1α表达,外源性HIF-1或内源性HIF-1α升高都能促进PAECs增殖和迁移,并且外源性HIF-1α会引起内源性HIF-1α表达增高,外源性HIF-1α转染成功的PAECs HIF-1α表达高于单纯缺氧。本研究结果还显示,HIF-1α+常氧组和HIF-1α+缺氧组PAECs的PDK1蛋白及mRNA表达量、乳酸及葡萄糖含量均高于常氧组和缺氧组,提示HIF-1α过表达可促进PDK1表达水平升高,增强PAECs的Warburg效应,促进增殖及迁移,这与Luo等 [ 29 ] 在胰腺癌小鼠模型中得出的结论相吻合。本研究在常氧条件下对PAECs进行HIF-1α外源性转染增强了PAECs的Warburg效应,与Chen等 [ 30 ] 的研究结果一致,同时,本研究发现PDK1表达水平上调,这与对PAECs进行单纯缺氧干预得出的结果一致,而2-ME可显著抑制这种变化,证明了缺氧PAECs异常增殖的作用机制可能为HIF-1α直接或间接对PDK1进行调控并影响Warburg效应。本研究应用HIF-1α特异性转录抑制剂2-ME后,2-ME+缺氧组各时间点HIF-1α及PDK1蛋白及mRNA表达水平均显著下降,Warburg效应减弱,缺氧诱导的细胞增殖和迁移得到有效抑制。因此,HIF-1α可通过调控PDK1影响Warburg效应,从而在PAECs增殖过程中起作用。而抑制HIF-1α表达可减少缺氧PAECs的PDK1表达水平,进而抑制其Warburg效应,对抗缺氧诱导的PAECs增殖。 目前临床上针对新生儿HPH的治疗大都局限于改善缺氧早期的肺血管痉挛,包括一氧化氮吸入、应用西地那非、米力农以及前列环素等 [ 31 ] ,一旦涉及到血管重塑阶段,则疗效欠佳。本研究结果显示,抑制HIF-1α或PDK1能够显著减弱缺氧环境下PAECs的Warburg效应,有效抑制PAECs增殖。因此,使用针对HIF-1α和PDK1的抑制剂可能减缓HPH气道重塑的病理进程,成为治疗新生儿HPH的重要手段,进一步探索HIF-1α和PDK1的上下游调控机制可以为临床治疗提供更为精准的靶向药物。此外,考虑到新生儿的特殊生理状态,开发的药物需要具有良好的耐受性和最小不良反应,以确保在不影响正常生理功能的前提下,有效控制HPH的进展。这些抑制剂在不同阶段HPH中的应用效果以及与其他治疗手段的联合应用也值得关注。 综上所述,本研究通过从细胞水平构建新生大鼠PAECs缺氧模型,发现对缺氧新生大鼠PAECs外源性转染HIF-1α可上调PDK1表达,增强缺氧新生大鼠PAECs的Warburg效应,促进其增殖和迁移,初步阐明了HIF-1α可能通过调节PDK1影响Warburg效应,参与了PAECs增殖的过程,为针对HPH气道重塑的治疗靶点提供了参考。但本研究也存在一些局限性,观察时间跨度较短,未分析缺氧程度的影响,HIF-1α与PDK1之间的具体上下游调控关系尚未完全明晰,后续仍需进一步研究,同时还应结合在体实验深入研究HIF-1α与PDK1的上下游调控机制,为临床治疗提供更精确的理论依据。 引用本文:罗洋,王乐,李珊珊,等. 缺氧诱导因子1α调控缺氧诱导的新生大鼠肺动脉内皮细胞增殖机制研究[J]. 中华新生儿科杂志(中英文),2025,40(04):226-233.DOI:10.3760/cma.j.cn101451-20240914-00326.



